Арийские народы на просторах Евразии. - Сибирь
- Просмотров: 864
Источник: А.А. Клёсов, К.А. Пензев. Арийские народы на просторах Евразии. (Серия «ДНК-генеалогия»).- М.: Книжный мир, 2015. - 352 с.
Легендарных Ариев нередко называют индоевропейцами. Но новейшие исследования в области ДНК-генеалогии показывают, что арийские народы шире индоевропейских языков. До времен, откуда пошли их предки, лингвистика, просто не дотягивается. Это носители гаплогруппы R1a, которые создали удивительные культуры и цивилизации по всей Евразии. Об этом рассказывается в новом незаурядном исследовании Анатолия Клёсова и Константина Пензева.
Арии вошли в историю и мифологию под разными именами, что маскировало их идентификацию. Это и «гипербореи» Русского Севера, и редины на Мальдивах тысячелетия назад, и «афанасьевцы» Минусинской котловины, и скифы от Дона до Алтая, и кельты Центральной Европы, и многие другие. Прямыми потомками древних Ариев являются многие миллионы славян. Сегодня термин «арий» в своем социальном значении открыт для людей всех рас, языков и национальностей. И авторы с удовольствием приглашают своих читателей обратиться к древней истории через призму новой науки - ДНК-генеалогии.
R1a, Абакан - Минусинск, Южная Сибирь, 3800-3400 и 2800-1900 л.н.
Еще одним значимым открытием является идентификация R1a в Хакасско-Минусинской котловине, за многие тысячи километров от Германии и от Русской равнины, с датировкой захоронения 3800-3400 л.н.[1], то есть тысячу лет спустя от R1a в Германии. Примерно 3600 л.н. арии ушли в Индию и Иран, точнее, в Индостан и на Иранское плато, и, как следует, проживали недалеко от Алтая и северной границы Монголии, в нынешних границах. Идентифицированные как R1a останки принадлежат к носителям андронов- ской археологической культуры.
Вот - эти ископаемые гаплотипы гаплогруппы R1a (3800-3400 л.н.):
13 25 16 11 11 14 10 14 11 32 15 14 20 12 16 11 23 (древние R1a).
В той же работе провели раскопки с датировкой 2800-1900 л.н., в захоронениях тагарской культуры, на той же территории, и обнаружили опять только гаплотипы группы R1a. Хотя прошли тысяча - две тысячи лет, гаплотипы отстались почти такими же:
13 24/25 16 11 11 14 10 13/1411 31 15 14 20 12/13 16 11 23 (древние R1a).

|
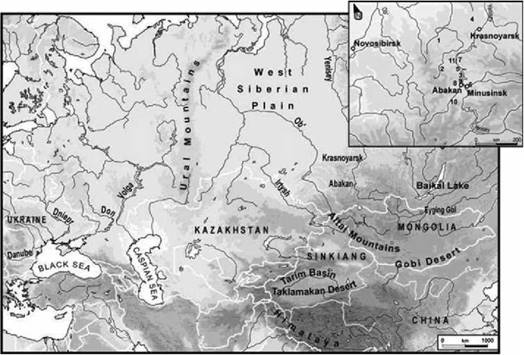
Рис. 21. На врезке показаны (нумерация) расположения археологических участков, из которых брали костные материалы для определения ДНК[2]. Видно, что это — далекое Зауралье - несколько тысяч километров на восток от Урала, к северу от монгольской границы, в приалтайском регионе. |
Найдены также некоторые объекты, у которых аллели начали чуть расходиться, но не у всех, и это несмотря на довольно большую временную дистанцию. В этом и состоит надежность гаплотипов - они меняются незначительно с течением времени. Если они изменились по нескольким маркерам - следовательно, прошли тысячелетия.
Кстати, необходимо еще отметить, что один га- плотип из десяти идентифицированных оказался из гаплогруппы С(хСЗ), т.е. гаплогруппа С, но не суб- клад СЗ. Нам это не очень интересно - и отнесение нечеткое, и гаплотип явно местный, никаких миграций не отражает. Потом к нему еще вернемся. Гаплотип его, понятно, совсем другой по сравнению с приведенными выше, почти все аллели разные:
14 22 15 9 12 13 11 14 12 30 16 14 19 11 15 10 22 [С(хСЗ)].
Пока мы его рассматривать не будем.
Смотрим выше на гаплотип из Германии и на га- плотипы этнических русских, для сравнения с ан- дроновскими гаплотипами из Минусинской котловины. Выделен маркер, который у древних гаплотипов в Германии не определялся, у русских гаплогруппы R1a он такой же - 11. Разница с гаплотипом общего предка этнических русских - только в паре 14-32 у ископаемых гаплотипов и 13-30 у предков этнических русских. Между ними две мутации, так как по правилам, детальные причины которых мы здесь объяснять не будем, это на самом деле пары 14-18 и 13-17. Числа 32 и 30 - это суммы первых двух, так принято представлять данные. Так вот. Пара 14-32 характерна для ариев Индии, Ирана, Ближнего Востока (ОАЭ, Бахрейн, Саудовская Аравия), то есть для собственно ариев, канонизированных, если так можно выразиться, лингвистической наукой и БСЭ. Расчетные датировки общих предков - 3500-4000 лет[3]. Вот - примеры современных гаплотипов их прямых потомков:
13 25 16 11 11 14 12 12 10 14 11 32-Индия (брамин)
13 25 15 11 11 14 12 12 10 14 11 32-Индия
13 25 15 10 11 14 12 13 10 14 11 32-Иран
13 25 16 11 11 13 12 12 11 14 11 32-ОАЭ
13 25 15 10 11 14 12 12 10 14 11 32-Араб (страна не указана)
13 25 15 10 11 14 12 12 10 14 11 32-Араб (страна не указана)
13 25 15 11 11 14 12 12 10 14 11 32-Бахрейн
13 24 15 10 11 14 12 12 10 14 11 32 - Саудовская Аравия
13 25 16 11 11 14 X X 10 14 11 32 - ископаемый гаплотип.
Так вот, все указанные выше гаплотипы Индии, Ирана, Ближнего Востока проанализированы весьма глубоко, денег эти люди, обладатели гаплотипов, не пожалели. Все они имеют цепочку характерных мутаций, снипов (от сокращения SNP), которая выстраивается так: R1a-L342.2-L657. Здесь присутствует уже не только общий род R1a, но и более конкретные племена- L342.2 (синоним - Z94). Очевидно это степные племена гапло- группы R1a, которые традиционно, в узком смысле, называются в археологии ариями. Арийским опять же традиционно в лингвистике считается их язык. Лингвисты и археологи не поясняют, как арийский язык, отраженный в виде топонимов, оказался представлен по всей Русской равнине, включая Русский Север, если арии обитали только в южных степях. Не поясняют они и то, как традиционный арийский знак - свастика - оказался опять же на всей Русской равнине, и как оказалось, что вологодские кружевницы веками, если не тысячелетиями, вплетают его в свои изделия. 184

|
Рис. 22. Вологодские кружева начала XX в. (цитировано в статье Klyosov A.A., Mironova Е.А. 2013)[4]. |
Не поясняют они и то, как «степной» арийский знак оказался на Балканах, в древней Адриатике (фото на рис. 23-29). Балканы, что хорошо известно, никак не относятся к южным русским и украинским степям. Короче говоря, лингвисты и археологи искусственно заужа- ют арийский ареал и диапазон арийских миграций. Для лингвистов - это только «иранцы» да «индоарии», но ДНК-генеалогия расширяет эти понятия.



|
Рис. 23-29. Надгробные памятники на Балканах, датировка неизвестна. |
Вся гаплогруппа R1a - это арийская гаплогруп- па, присутствие «степных ариев» в Европе является только лишь фрагментом жизни и миграций древнего народа, который прибыл в Европу, на Балканы не менее 8 тыс. л.н., а скорее 9-10 тыс. л.н., и оттуда, после распространения по всей Европе, двинулся на Русскую равнину, о которой у ариев остались чудесные воспоминания как «Ариана Ваэджа», «арийский простор».
Весьма вероятно, что племя L342.2 сначала прибыло с Балкан в Днестро-Донской регион, а потом двинулось далее на восток по южному, степному и лесостепному пути. Затем, весьма вскоре, у кого-то из ариев произошла снип-мутация L657, потомки выжили и приумножились, и вот мы видим тот же снип, метку степных ариев, и на Ближнем Востоке, и в Индии, и в Минусинской котловине в Южной Сибири.
Вот так определение ископаемых гаплотипов продвигает наши знания, конечно же в совокупности с обширными данными ДНК-генеалогии и других дисциплин.
Гаплогруппа R, Южная Сибирь, 24 тыс. л.н.
Археологические раскопки в селении Мальта, в 86 км к северо-западу от Иркутска, на реке Белой, велись с 1928 г. Здесь найдены тысячи древних арти- фактов, и что наиболее важно - двойное захоронение детей. Датировки (радиоуглеродные) обычно варьировались от 19,900±800 до 21,700±160 л.н. В работе Raghavan М. и др. (2013)[5] они были тщательно переделаны, и оказались в интервале 24.423-23.891 л.н., то есть примерно 24 тысячи л.н.
Y-хромосомная гаплогруппа была идентифицирована как R, митохондриальная как U[6].
Авторы заключили, что по геномным показателям древний человек из Мальты был близок к американским индейцам, и пресса подала это заключение как очередную сенсацию, чего не избежала и научная литература[7]. На самом деле авторы, не слишком знакомые с ДНК-генеалогией, не подумали, что гаплогруп- пы R и Q - братские (см. рис. 12), они образовались от общего предка - гаплогруппы Р, и, естественно, их геномы должны быть ближе других. Авторы так и сообщили, что ископаемый геном ближе всего к американским индейцам (имеющим в основном га- плогруппу Q, которой в Европе почти нет (обычно от О до 0,5%, в России в среднем 1,5% - данной информации в статье авторов нет), и стали фантазировать о том, что те, в Мальте, предки американских индейцев. Пресса, естественно, подхватила сенсацию, это приняли и в современной популяционной генетике и в СМИ.
Несмотря на грубые ошибки в интерпретации данных, сами данные очень важны, и могут стать дополнительным обоснованием для утверждения о появлении гаплогрупп R1a и Rib в Южной Сибири[8].
Причины монголоидности восточных скифов
Пазырыкская культура - это археологическая культура железного века (III—V вв. до н.э., хотя некоторые исследователи опускают дату до VI в. до н.э.), которую относят к «восточному скифскому кругу». Регион ее распространения включает в себя горный Алтай и смежные территории Казахстана и Монголии.
Недавно изучались гаплотипы и гаплогруппы (мужские и женские) современных жителей этого региона[9], и митохондриальные гаплогруппы (женские по сути, поскольку мужчины получают их от матери, но дальше не передают, нет митохондрий в сперматозоидах) ископаемых костных остатков пазырыкской культуры[10]. Оказалось, что большинство мужских гаплогрупп в регионе относятся к гаплогруппе R1a, причем наибольшее их количество было в народности алтай-кижи. R1a содержали также тубалары, чел- каны, кумандины. На втором месте по численности была гаплогруппа Q, затем С, потом N, остальные - минорные, единичные гаплогруппы, включая Rib, которые, как правило, случайны, и могли попасть туда в любое время.
У носителей гаплогруппы R1a на Алтае была характерная особенность. Если на Русской равнине и в Центральной Европе у носителей R1a присутствует преимущественно митохондриальная гаплогруппа (мтДНК) Н, как и у их жен и подруг, так называемая «европейская», или «западная» мтДНК, то на Алтае у R1a, в основном, восточные, «восточно-евразийские», азиатские мтДНК - А, С, D и G, их у носителей гаплогруппы R1a до половины и двух третей, остальные - минорные, единичные. Европеоидных мтДНК у них почти нет.
Носители мтДНК А, С, D и G, как правило - монголоидные женщины и их сыновья и дочери, опять же монголоидные. Вот в этом и состоит ответ на вопрос, почему алтайские носители гаплогруппы R1a, потомки ариев-скифов, как правило, сами монголоидны. Антропологию в значительной степени определяют женщины. К тому же, у алтайских R1a те же монголоидные женщины (в процессе воспитания малолетних детей), видимо, сменили язык всей популяции на тюркский.
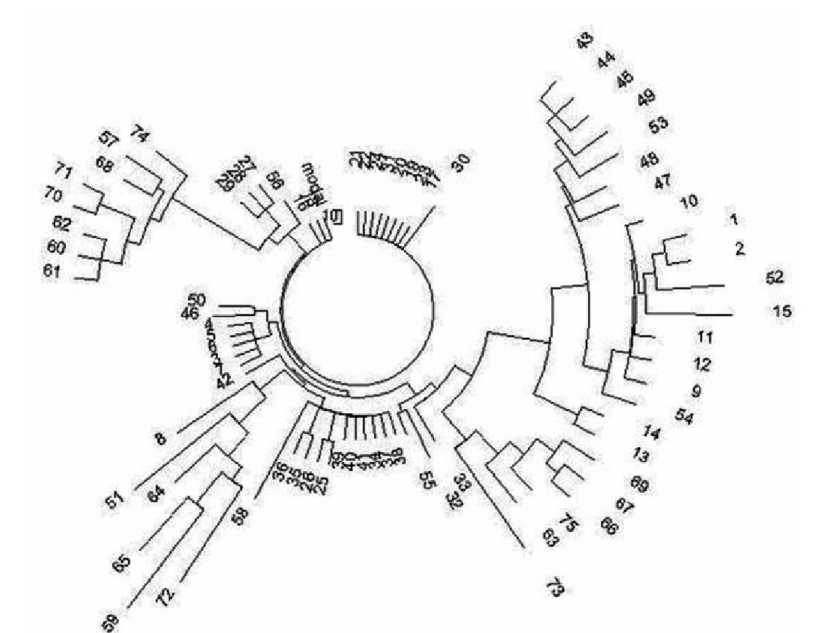
Анализ гаплотипов алтайцев с точки зрения ДНК-генеалогии проведен в статье А.А. Клёсова (2012)[11]. Дерево гаплотипов (рис. 39) имеет необычный вид:
|
Рис. 39. Дерево из 75 гаплотипов гаплогруппы R1a на Алтае в 17-маркерном формате. Верхняя левая ветвь - тубала- ры (57, 68, 70, 71, 74), челканы (60, 61, 62) и алтай-кижи (57). Нижняя правая ветвь - подобный состав: тубалары (66, 67, 69, 73, 75) и один челкан (63). Нижняя левая ветвь — смешанная: кумандины (64, 65), тубалар (72), алтай-кижи (8, 51, 59). Остальные гаплотипы - алтай-кижи. |

Вид необычный потому, что гаплогруппа одна, то есть род один, а ветви расходятся в разные стороны, будучи заметно изолированными друг от друга. Это означает, что у данной популяции была тяжелая судьба. Племена погибали, немногие спасшиеся бежали, начинали свои генеалогические линии практически с нуля. Это повторялось, и опять спасались, бежали, и опять начинали свои линии. То есть повторялся эффект «последнего из могикан». В итоге все ветви на дереве гаплотипов относительно молодые, это набор относительно молодых «кустов», пересаженных со старых черенков, которые погибли. Но исходный общий предок жил в далекой древности, на это указывают масштабные расхождения между ветвями.
Для сравнения, дерево гаплотипов группы R1a на Русской равнине выглядит несравненно более благополучно: [12]
Еще более благополучно выглядит дерево гапло- типов Русской равнины для большого числа (801) протяженных гаплотипов (67-маркерных)[13].
Посмотрим более внимательно на современные алтайские гаплотипы. Левая верхняя ветвь на рис. 39 из восьми гаплотипов (тубалары и челканы) имеет следующий предковый гаплотип:
13 24 16 9 12 14 10 14 11 32 14 14 20 12 17 11 23 (алтайские гаплотипы, ветвь).
В ветви всего 10 мутаций на 8 гаплотипов, то есть на 8x17 = 136 маркеров, что дает 10/8/0.034 = 37 —► 38 условных поколений, то есть 950±315 лет до общего предка (стрелка - поправка на возвратные мутации, 0.034 - константа скорости мутации для 17-маркерных гаплотипов). Иначе говоря, общий предок этой ветви жил примерно в XI в. нашей эры, плюс-минус три века. Ясно, что он удален от скифов по времени, но гаплотип показывает, что современные алтайцы - прямые предки скифов. Тип гаплотипа тот же, та же самая пара 14-32.
13 25 16 11 11 14 1014 11 32 15 14 20 12 16 11 23 (ископаемые скифы, R1a).
У подножия описанной ветви находится миниветвь из четырех гаплотипов популяции алтай-кижи, три из которых одинаковы, а четвертый (56) отличается всего на одну мутацию:
13 26 16 10 11 14 10 14 11 32 15 14 21 12 16 11 23 (№ 27, 28,29)
13 25 16 10 11 14 10 14 11 32 15 14 21 12 16 11 23 (№ 56).
Мы видим, что у них та же самая характерная «подпись» — пара 14-32. Более того, у них всего три и две мутации, соответственно, от ископаемого скифского гаплотипа, то есть разницы почти нет.
Одна мутация между приведенными выше гапло- типами помещает их мини-ветвь на 1/0.034 = 29 —> 30 поколений, то есть примерно на 750 л.н. Но между этой мини-ветвью (алтай-кижи) и ветвью тубаларов и челканов - 6.5 мутаций, или 5900 лет между их общими предками, что помещает их общего предка на (5900+950+750)/2 = 3800 л.н. Это как раз датировка ископаемых гаплотипов скифов.
Иначе говоря, от ДНК-линии, идущей от 3800 л.н. (которая, в свою очередь, тоже явно прошла бутылочное горлышко популяции) остались две подветви с возрастом 950 и 750 л.н. Но расстояние между ними выдает, что они далеко разошлись от их общего предка, и на сколько они разошлись друг от друга можно без труда вычислить. Это расстояние между общими предками двух алтайских ветвей и составляет указанные выше 5900 лет.
Гаплотипы, подобные древним скифским, имеют и остальные ветви дерева гаплотипов на рис. 39. Например, малая ветвь слева из семи гаплотипов (в которых всего три мутации):
13 25 16 11 11 14 10 14 11 32 - 15 14 21 10 16 11 23 (325 лет до общего предка).
Довольно древняя ветвь из 6 гаплотипов (на 7 часов) [27 мутаций]:
13 25 15 10 11 14/15 10 13 11 30/31 - 15 14 20 12 16 11 23 (3800 лет до предка).
Это вполне может быть субклад L342.2, базовый гаплотип которого на западе Русской равнины выглядит следующим образом:
13 25 16 11 11 14 10 13 11 30 - 15 14 20 12 16 11 23 (L342.2 на западе).
Молодая ветвь из 10 гаплотипов внизу дерева, в которой всего 4 мутации:
13 25 16 11 11 14 10 14 11 32 - 15 14 21 12 17 11 23 (300 лет до общего предка).
Ветвь из семи гаплотипов в верхней правой части дерева, в которой всего 5 мутаций, что дает 5/7/0.034 = 21 поколение, или примерно 525 лет до общего предка:
13 26 16 10 11 17 11 14 11 32-15 14 19 11 15 11 23.
Подветвь из 9 гаплотипов на 3 часа (в которой всего 7 мутаций, то есть общий предок жил 7/9/0.034 = 23 поколения, то есть 575 л.н.), с базовым гаплотипом:
13 26 16 11 11 17 11 14 11 31 - 15 14 19 11 15 11 23.
Видно, что это родственная ветвь предыдущей. У них одинаковые значения большинства аллелей, и они отличаются всего двумя мутациями, то есть их общие предкирасходятся на2/0.034= 59—>63 поколения, то есть на 1575 лет. Их общий предок жил (1575+525+575)/2 = 1340 л.н. Ясно, что эта двойная ветвь - молодая (в отношении своего общего предка). Видно, как фрагментированы ветви, как они рассыпаются на совсем недавно выжившие и давшие недавних же потомков.
Основной вывод таков - все эти современные алтайские гаплотипы, точнее, их носители, являются потомками древних скифов, тех же древних ариев, с предком на Русской равнине.
Раскопки пазырыкской культуры выявили три мтДНК, относящиеся к бронзовому веку и шестнадцать мтДНК - к железному веку. К сожалению, Y-хромосомные ДНК не изучались, тем не менее полученная информация оказалась важной. Из всех 19 ископаемых мтДНК гаплогрупп 11 оказались азиатскими (А, С, D, и G), и 8 - западными, точнее, западно-евразийскими (HV, J, U, Т, К). Все три гаплогруппы бронзового века оказались азиатскими. Гаплогруппы железного века дали смесь европейских и азиатских гаплогрупп. Это в любом случае показывает, что скифы были по антропологии и европеоидные, и монголоидные, причем арии-скифы, пришедшие в алтайский регион в бронзовом веке, то есть самые ранние, брали в жены местных монголоидных женщин, и их потомки, сохраняя гаплогруппу R1a, были уже монголоидными. Последнее объясняет монголо- идность некоторых скифов, кочевавших по степям Евразии. Многие скифы, очевидно, откочевывали на восток со своими европейскими женами и подругами, что и дало «западные» мтДНК в их потомках, в том числе и на Алтае в древности. Отсюда идет разнообразие антропологии скифов-кочевников, от европео- идности до монголоидности, при наличии основной гаплогруппы R1a.
Итак, в отношении происхождения родов и племен, гаплогрупп и субкладов в ДНК-генеалогии, понятия ариев, скифов, восточных славян взаимосвязаны и взаимозаменяемы, мы просто относим их к разным временным периодам, в соответствии с принятой в исторической науке терминологией. Реальность состоит в том, что древние арии населявшие Русскую равнину и были теми самыми прото- и праславянами, которых, упорно ищут, но как в заколдованном лесу возвращаются ко временам Иордана.
Что касается лингвистики, то один из авторов этой книги однажды выдвинул предположение, названное им арийской теоремой, суть которой состоит в следующем.
Первичная арийская общность (индоевропейский унитарный пранарод), образовавшаяся в результате распадения ностратической общности, не делилась в первичном ареале своего зарождения (Северо-Восточная Европа) на лингвистические разделы иранцев, германцев, кельтов и пр. Только много позже, к началу Средневековья, она стала разделяться на ряд этносов именуемых, впоследствие и по совокупности, общим термином славяне.
Механизм же образования иных индоевропейских этнолингвистических общностей, как-то иранцы, индоарии, греки, латиняне и пр. следующий. Миграции ариев (т.е. представителей индоевропейского унитарного пранарода) на восток в Сибирь, Дальний Восток, Центральную Азию; на юг в Малую Азию, Иран, Индию; на запад в Западную Европу, приводили к завоеванию местных неиндоевропейских народов и образованию новых этносов, в которых арии представляли из себя господствующий социальный слой. Таким образом и появились более поздние индоевропейцы как ответвления от главного арийского ствола.
Картина, которая здесь, в теореме, представлена, является, конечно же, схематичной и очень упрощенной, но, как мы понимаем, она дает представление о разделении индоевропейских народов на две группы: 1-я группа, этносы возникшие в результате распада первичной арийской общности, их языки развивались, более или менее, естественным путем (к примеру, сегодняшние великороссы, белорусы, малороссы и др.); 2-я группа, этносы возникшие в результате арийских завоеваний или мирной инфильтрации, их языки, исконно неарийские, претерпели изменения под арийским влиянием (к примеру, большинство сегодняшних европейцев, индийцев, иранцев и др.).
Данная теорема справедлива в отношении времен, последовавших в Северо-Восточной Европе вслед за Неолитической революцией, когда у ариев появилось производящее хозяйство, произошла социальная дифференциация и образовались три основных арийских касты: жреческая, воинская и торгово-ремесленная.
Таким образом, если не считать лингвистический фактор определяющим, а только одним из целого ряда этнических, культурных, профессионально-социальных, хозяйственно-бытовых и прочих факторов, и учитывать категорию родства (очень важную во все времена и при любых обстоятельствах), то проявляется вполне четкая картина: арии, скифы и восточные славяне - это одни и те же люди, в своей естественной хронологической динамике. Большинство их принадлежало и принадлежит одному и тому же роду - R1a.
Несомненно, что в составе арийских военных, духовных и торгово-ремесленных корпораций на раннем этапе были представлены в основном носители гаплогруппы R1a, которые со временем разбавлялись людьми с самыми разнообразными гаплогруппами и самого разного происхождения, поскольку основным критерием подбора личного состава в подобных корпорациях являлся высокий профессионализм. Это привело к тому, что даже в высших кастах Индии га- плогруппа R1a, исходно арийская, достигает максимальной доли в 72%, а в некоторых регионах падает до 50%, и даже 40% (в провинциальных регионах Индии). Впрочем, для этого понадобилось более трех тысяч лет. Сейчас подобные разбавления происходят значительно быстрее, особенно в странах Запада. Например, конгресс США за 225 лет своего существования (с 1789 г.) с почти чисто англо-саксонского состава стал по сути полиэтническим. Чтобы не быть голословными, отметим, что в Конгрессе США созыва 2013-2014 гг. среди конгрессменов присутствуют 44 афроамериканца, 37 «латино», 13 «азиатов по происхождению» и тихоокеанских островитян, и два американских индейца. Поэтому термин «арий» в своем социальном значении открыт для людей всех рас, языков и национальностей.
[1] Ancient DNA provides new insight into the history of south Siberian Kurgan people / Keyser C. [et al.] // Hum. Genet., 2009. Vol. 126. P. 395-410.
[2] Ibid
[3] КлёсовА.А. Субклад R1a-L342-L657 в древнем Зауралье // Вестник Академии ДНК-генеалогии, 2013. Т. 6. № 3. С. 446-451.
[4] Klyosov A.A., Mironova Е.А. A DNA Genealogy Solution to the Puzzle of Ancient Look-Alike Ceramics across the World// Advances in Anthropology. 2013. Vol. 3. No.. P. 164-172.
[5] Upper Palaeolithic Siberian genome reveals dual ancestry of Native
Americans / Raghavan M. [et al.] // Nature, 2013. doi:10.1038/
naturel2736
[6] Ibid.
[7] Balter М. Ancient DNA links Native Americans with Europe // Science. 2013. Vol. 342. P. 409-410.
[8] Klyosov A.A. DNA Genealogy, mutation rates... P. 217-256; Klyosov A.A., Rozhanskii I.L. Haplogroup R1a as the Proto Indo-Europeans and the legendary Aryans as witnessed by the DNA of their current descendants// Advances in Anthropology. 2012. Vol. 2. No.l. P. 1-13; Klyosov A.A. Ancient history of the Arbins... P. 87-105.
[9] Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and indigenous Altaians / Dulik M.C. [et al.] // Amer. J. Human Genetics. 2012. 90, 1-18. DOI 10.1016/ajhg.2011.12.014
[10] Tracing the origin of the East-West population admixture in the Altai region (Central Asia) / Gonzalez-Ruiz M. [et al.] // PLOS One. 2012. Vol. 7. P. 1-11. e48904
[11] Клёсов А.А. Гаплотипы группы R1a на Алтае: «автохтоны» и «индоевропейцы» // Вестник Академии ДНК-генеалогии. 2012. Т. 5. №12. Р. 1511-1525.
[12] Клёсов А.А. Гаплотипы восточных славян: девять племен? // Вестник Российской Академии ДНК-генеалогии. 2009. №2. Т. 2. С. 232-251.
[13] Rozhanskii I.L., Klyosov А.А. Haplogroup R1a, its subclades and branches in Europe during the last 9000 years // Adv. Anthropol., 2012. Vol. 2. No. 3. P. 139-156.